
RAD52 Primary Antibody
Item Information
Catalog #
Size
Price
Description
The protein encoded by this gene shares similarity with Saccharomyces cerevisiae Rad52, a protein important for DNA double-strand break repair and homologous recombination. This gene product was shown to bind single-stranded DNA ends, and mediate the DNA-DNA interaction necessary for the annealing of complementary DNA strands. It was also found to interact with DNA recombination protein RAD51, which suggested its role in RAD51 related DNA recombination and repair. A pseudogene of this gene is present on chromosome 2. Alternative splicing results in multiple transcript variants. Additional alternatively spliced transcript variants of this gene have been described, but their full-length nature is not known.
Product Overview
Entrez GenelD
5893
Aliases
N
Clone#
5E11E7
Host / Isotype
Mouse / IgG1
Species Reactivity
Human
Immunogen
Purified recombinant fragment of human RAD52 (AA: 269-418) expressed in E. Coli.
Formulation
Purified antibody in PBS with 0.05% sodium azide
Storage
Store at 4°C short term. Aliquot and store at -20°C long term. Avoid freeze/thaw cycles.
Product Applications
WB (Western Blot)
1/500 - 1/2000
ICC (Immunocytochemistry)
1/200 - 1/1000
FCM (Flow Cytometry)
1/200 - 1/400
ELISA
1/10000
References
1.Blood. 2013 Aug 15;122(7):1293-304.
2.PLoS One. 2012;7(11):e50461.
2.PLoS One. 2012;7(11):e50461.
Product Image
Elisa

Figure 1: Black line: Control Antigen (100 ng); Purple line: Antigen(10ng); Blue line: Antigen (50 ng); Red line: Antigen (100 ng);
Western Blot
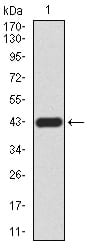
Figure 2:Western blot analysis using RAD52 mAb against human RAD52 (AA: 269-418) recombinant protein. (Expected MW is 42.5 kDa)
Western Blot

Figure 3:Western blot analysis using RAD52 mAb against HEK293 (1) and RAD52 (AA: 269-418)-hIgGFc transfected HEK293 (2) cell lysate.
Western Blot
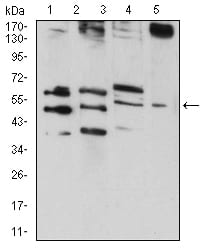
Figure 4:Western blot analysis using RAD52 mouse mAb against HepG2 (1), MCF-7 (2), MCF-7 (3), and C6 (4) cell lysate.
Immunofluorescence analysis

Figure 5:Immunofluorescence analysis of HeLa cells using RAD52 mouse mAb (green). Blue: DRAQ5 fluorescent DNA dye. Red: Actin filaments have been labeled with Alexa Fluor- 555 phalloidin. Secondary antibody from Fisher (Cat#: 35503)
Flow cytometric

Figure 6:Flow cytometric analysis of MCF7 cells using RAD52 mouse mAb (green) and negative control (red).
For Research Use Only. Not for use in diagnostic procedures.